摘要:建立预测动物生长的数学模型时,模拟氨基酸的吸收代谢具有特别重要的意义,因为预计的蛋白质沉积速率与预期的体重间往往不成比例。事实上,氨基酸的吸收和代谢是一个复杂的过程,并且与机体细胞内及细胞间物质的连续流动高度整合在一起。为了建立氨基酸的利用模型,必需构建一个用离散的生理和代谢过程进行描述的简化的氨基酸代谢框架。并将以上离散的氨基酸代谢过程分为维持相关过程和生长相关过程。在此,生长被看作是机体营养物质沉积及与此直接相关的氨基酸代谢过程的函数。本文中将主要介绍体蛋白质的沉积、蛋白质的消化、氨基酸的必然分解代谢、氨基酸的优先分解代谢。用数学模型表述动物的生长过程已经成为一个主流,并且数学模型在研究、教学及商业实践中的应用也日益增加。当这些模型的建立更多的以因果关系而不是以严格的试验经验为基础时,模型的有效性和实用性将进一步提高。
关键词:氨基酸;模型;猪;蛋白质;利用率
1. 前言
准确测定动物对营养物质的需要量是动物营养学中一项非常基础的工作。 在过去的几十年中,我们在评定动物的营养需要量方面已经做了大量的工作,并且对评定动物营养需要量的理论及方法的认识也在不断深入。 传统的基于“剂量-效应”关系的研究为动物营养需要量的评定提供了大量初级的数据,这为提出更灵活、更完善的预测动物营养需要量的方法铺平了道路。
动物对蛋白质的营养需要在本质是对氨基酸的需要。 尽管很多试验研究已经给出了猪氨基酸需要量的推荐值,但是这些值之间还是存在着明显的差异。 绝大多数估计氨基酸需要量的方法是“剂量-效应”法。 这一方法的关键是配制一种基础饲粮,这种基础饲粮必需保证除被考查的氨基酸外其它养分必需充足。 然后向这一基础饲粮中添加不同梯度的限制性氨基酸,测定所选指标的变化情况,最后确定这一限制性氨基酸的需要量。 这种方法存在着明显的不足。 首先,为了确保其它氨基酸不缺乏,必需超量添加。 如此一来,则会造成氨基酸的不平衡,从而会影响动物的生长速度和降低动物的随意采食量。 其次,没有统一的效应指标。 此外,得出“剂量-效应”曲线后,不同的求解标准也会造成氨基酸需要量评定的差异。
由于传统试验评定方法的固有缺陷,建立适用范围更广的因果模型以模拟饲粮氨基酸的吸收利用情况已经引起人们的关注。 早期的析因模型就是一种简单的静态因果模型。 当运用析因法建立起动态的模型时,我们则可以估计出动物任意生产阶段和生产状态下的氨基酸需要量。 当模型建立在更为通用的原则基础之上时,影响氨基酸需要估计准确度的只有模型本身,因而这种模型可以运用到具有不同生产潜能的其它群体中。 更为重要的是,这种模型不仅可以估计营养物质的需要量,而且还能让我们更深入的认识与这种需要的产生有关的代谢过程的生物学机制。
2. 氨基酸利用率的模拟
从粪尿中排出的氮的总量约占猪从饲粮中摄入的氮总量的2/3,其中1/4以上的氮流失是由于饲粮中蛋白质的质量和数量没有与动物生长的需要量正确匹配,不能发挥动物的最佳生产性能和饲粮利用效率所致(Whittemore等,2001a,2001b)。 Moughan等(2003)对饲粮粗蛋白的利用率进行预测,发现粗蛋白的利用率均远低于50%(表1)。 表 1给出了50 kg小母猪在不同采食量下对6种商品生长猪料中粗蛋白和赖氨酸的利用率。 可见,在低采食量的情况下,饲粮粗蛋白的利用率为20-42%,而在高采食量的情况下为22-35%。 平均来看,饲粮粗蛋白的利用率接近30%,大约70%的氮被猪排出体外。 饲粮氨基酸的不平衡是引起饲粮中蛋白质利用率降低的原因之一。 在表 1的六种饲粮中,赖氨酸为第一限制性氨基酸,饲粮氨基酸的不平衡性对其利用率不构成影响,因此对饲粮赖氨酸的利用率也进行了预测(Moughan等,2003)。 结果发现,赖氨酸的利用率高于粗蛋白,摄入的赖氨酸的利用率平均接近44%,但是,仍然有一半以上的赖氨酸没有用于机体瘦肉组织的沉积。 从这些数据可以看出,充分认识导致机体氨基酸流失的生理过程,对解释饲粮限制性氨基酸用于生长沉积的低效性非常重要。
动物对氨基酸的吸收和代谢是一个复杂的过程,与机体细胞内和细胞间连续的物质流量密不可分。 为了建立氨基酸的利用模型,必需构建一个用离散的生理代谢过程进行描述的简化的氨基酸代谢框架(表 2)。 这些过程可分为氨基酸吸收前过程和氨基酸吸收后过程两类。 氨基酸吸收前过程包括动物将氨基酸摄入消化道及随后氨基酸的消化吸收过程。 氨基酸吸收后过程又分为维持相关的代谢过程和生长相关的代谢过程(Moughan等,2003)。 在这里,生长被视为养分沉积,以及与养分沉积直接相关的物质消耗的函数。

Moughan等(2003; 2008)通过模拟的数据(表 4、表 5),给出了各氨基酸代谢过程的相对重要性。 表 4中给出了体重为50kg生长猪在饲喂商品饲粮(表3)时,采食量和最大蛋白质沉积速率(Pdmax)对赖氨酸利用的影响。 在该模型中,采食量对维持需要下尿氮的损失没有影响,而Pdmax仅对其有微小的影响。 因氨基酸的必然分解代谢(inevitableamino acid catabolism)引起的尿氮损失随着采食量的提高而增加,在给定的采食量下,通常随着Pdmax的增加而下降。 但是在采食量高的情况下氨基酸必然分解代谢引起的尿氮损失保持不变。 氨基酸过量引起的尿氮损失在采食量高时据有重要意义,而氨基酸的优先分解代谢(preferentialamino acid catabolism)造成的尿氮损失则刚好相反。 采食量和Pdmax均不影响皮屑氨基酸损失量。 肠道内源氨基酸的损失受采食量的影响,而不受Pdmax的影响。 由于赖氨酸是该商品饲粮中的第一限制性氨基酸,因此,氨基酸不平衡造成的赖氨酸损失量为几乎为零。

表4中的数据表明,氨基酸的必然分解代谢(inevitable amino acid catabolism)可能对第一限制性氨基酸的利用率有重要的影响,特别是在高采食量的情况下。 未吸收的氨基酸及肠道内源氨基酸损失也占有重要的比重。 体蛋白质周转引起的氮损失所占比重相对较小,而皮屑引起的氨基酸损失数量更低。 当饲粮代谢能含量成为体蛋白质沉积的限制因素时,氨基酸的优先分解代谢可能占据氨基酸损失的相当大的比重。 因而,供给过量的氨基酸也会引起氨基酸利用率降低。 而表 5中的数据则清楚的表明,对于配制合理的商品饲粮,因氨基酸不平衡性引起的氨基酸损失很小,所以饲粮氨基酸的不平衡性对这种饲粮中氨基酸利用率的影响相对而言并不大。
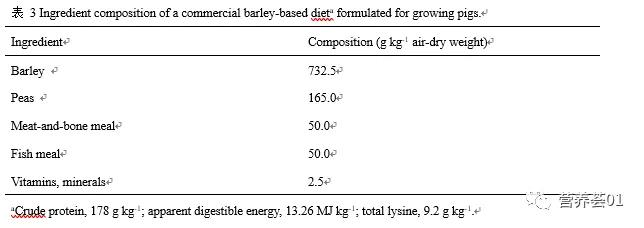
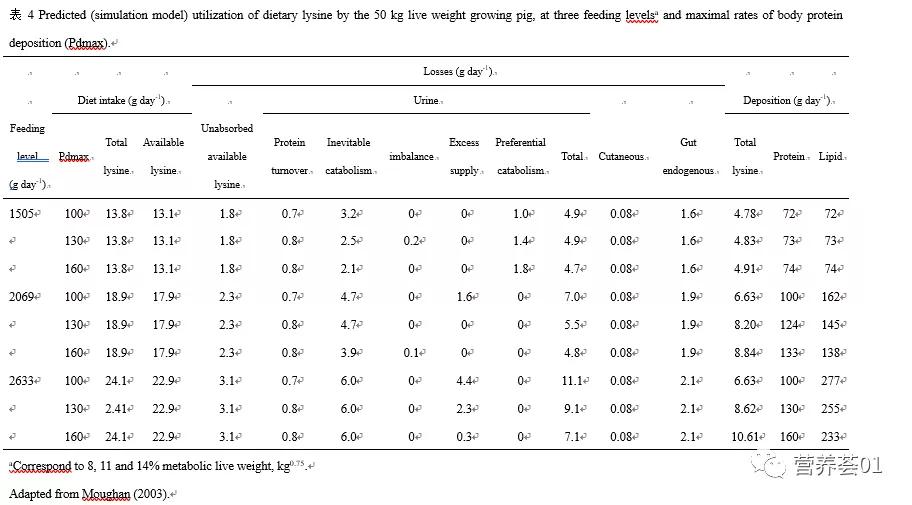
从以上数据可以看出,蛋白质的沉积、蛋白质消化、氨基酸的必然分解代谢(inevitableamino acid catabolism)、氨基酸的优先分解代谢及维持相关的代谢过程对机体整体蛋白质的代谢有着重要的意义。 而这些代谢过程对机体整体蛋白质代谢的影响因饲粮类型、动物年龄及体重的改变而改变。 尽管如此,运用以上简化的模型还是可以让我们概略地了解氨基酸代谢的动力学过程,并对各个代谢过程的重要性进行排序定位。 下面将介绍几个对生长有重要影响的氨基酸代谢过程。
3. 蛋白质的沉积
生长猪每日的氨基酸需要中,约90%以上用于生长相关的过程,其中用于满足新生组织沉积的氨基酸需要占有重要的分量。 不过,在新蛋白质合成的过程中,一些辅助过程消耗的氨基酸在数量上也占有较大的比重,如氨基酸的必然分解代谢(inevitable amino acid catabolism)、肠道内源氨基酸损失、新蛋白质合成相关的体蛋白周转、非氨基酸和非蛋白质含氮物的合成,以及氨基酸的优先分解代谢。 因而在构建氨基酸利用率的模型时需要考虑这些代谢过程。
为了预测每日氨基酸沉积量,必需首先预测出每日蛋白沉积率(daily rate of protein retention),然后结合体蛋白质氨基酸组成(理想氨基酸模式)则可计算出新合成蛋白质中的氨基酸沉积量。 可是,对于生长快速的动物而言,胴体和内脏蛋白质的氨基酸组成并不相同,胴体蛋白质日沉积量和内脏蛋白质日沉积量也不相等。 因而有必要对这两个部分氨基酸的需要量分别进行估计,然后对这两部分氨基酸的沉积速率进行加权后得出总的理想氨基酸模式。
细胞每天合成蛋白质的数量是有限的,因而机体蛋白质的日沉积速率有一个固有的最大值(Pdmax)。 当摄入的氨基酸(即使是理想氨基酸)超过动物机体蛋白质合成能力的上限时,多余的氨基酸将被分解代谢。 Pdmax受动物基因型、性别和年龄的影响,其平均值可低至90 g/d,最高在200 g/d以上(Whittemore,1983)。 年龄对Pdmax的影响还需要进一步的研究,人们就这一问题提出了不同的数学模型(Moughan,1999; Whittemore等,2001)。 通常简单的认为,在动物的生长期内Pdmax保持不变。 对于猪,以上假设在体重相对较低时可能是合理的近似,但是当体重超过这一范围或对于生长速度更快的动物而言可能不再正确。 Emmans(1986)用曲线模型模拟动物生长: 从动物出生开始,Pdmax逐渐增加,直到最大值,随后开始降低,直至体成熟,此时Pdmax等于零。
对于猪的生长模型而言,Pdmax作为一个重要的限制条件已经被研究者采用,而描述Pdmax随动物年龄变化的更复杂的方法也已经有人提出(Wellock等,2004)。
4. 蛋白质的消化
4.1氨基酸的生物学利用率(amino acidbioavailability)
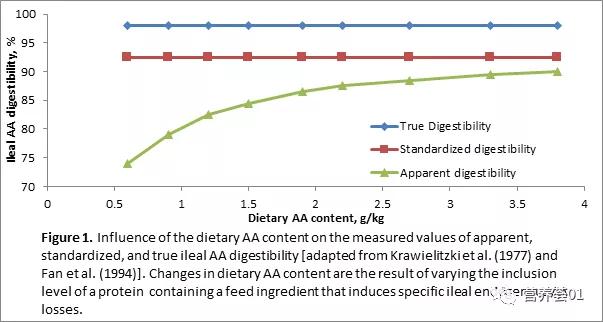
准确测定饲粮中每种必需氨基酸的生物学利用率是评价原料营养价值及估计动物氨基酸需要量的关键。 氨基酸消化率(amino acid digestibility)是评价氨基酸生物利用率(aminoacid bioavailability)的一种方式。 自20世纪70年代中期以来,在评定饲粮氨基酸消化率及预测氨基酸每日吸收量方面已经取得了很大的进步。 对氨基酸消化率的评定已经从不准确的肛门收粪测定粗蛋白的消化率进步到标准化回肠可消化氨基酸(standardized ileal aminoacid digestibility,SID)(Stein等,2007)。 SID可通过测定回肠末端末吸收的氨基酸含量,并用内源性氨基酸含量进行校正后求出。 回肠末端内源性氨基酸损失可分为两个主要部分: 基础损失(basal losse)和特异性损失(specificlosses)(Stein等,2007)。 基础损失,以前称为非特异性损失或非饲粮依赖性损失,表示动物机体氨基酸的最小必然损失量。 通常认为这一部分损失与消化道内饲粮干物质流量或动物的代谢状态有关,而与饲粮组成无关。 特异性损失受饲粮组成成分的影响,以前称为额外损失或饲粮依赖性损失,指由饲粮中特定组成成分(如养分含量、纤维及抗营养因子的类型等)引起的超出基础损失部分的氨基酸损失。 当饲喂消化率较高的纯化蛋白质(如酪蛋白或卵蛋白)时,特异性损失最低。 相反,如果饲粮中含有纤维素或抗营养因子,特异性损失可能占总回肠内源氨基酸损失(IAAend)的50%以上。 因而,回肠末端食糜中的氨基酸可分为3部分: 饲粮中未消化氨基酸,基础回肠内源氨基酸(basal IAAend)和特异性回肠内源氨基酸(specific IAAend)。 根据回肠末端食糜中氨基酸测定方法的不同,饲粮氨基酸的消化率可分为: 表观回肠可消化氨基酸(apparent ileal aminoacid digestibility,AID)、标准化回肠可消化氨基酸(standardized ileal amino acid digestibility,SID)和回肠真可消化氨基酸(trueileal digestibility,TID)。 三者之间的关系见figure1 。 计算公式分别如下:

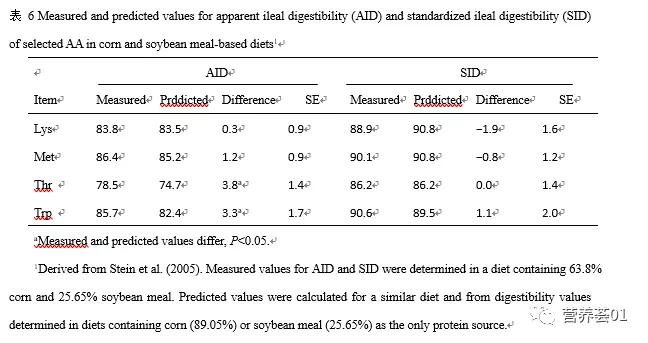
与AID相比,使用SID的主要优点再于SID的数值在配合饲粮时更具有可加性(Stein等,2005,表 6 ),能够较准确的预测不同原料配合时饲粮中氨基酸的消化率。 因此,在实际配制饲粮或解释试验现象时,使用SID可以克服使用AID和TID的一些不足和限制。 大多数猪饲料原料的SID已经给出(NRC,1998)。
近年来,有关消化道上段微生物是否为宿主净合成必需氨基酸的问题受到关注。 目前有一点是肯定的,即肠道微生物确实要合成饲粮中的必需氨基酸,如赖氨酸(Fuller和Tome,2005)。 Torrallardona等(2003)利用15NH4Cl和14C-Polyglucose标记进行试验,以估计20kg仔猪肠道中微生物的氨基酸合成及随后在回肠可消化率。 结果表明,吸收的氨基酸中,由微生物合成的赖氨酸为1.1 g/d,亮氨酸为2.0g/d,异亮氨酸为0.8 g/d,苯丙氨酸为0.3g/d,缬氨酸为1.8 g/d。 这些由微生物合成的氨基酸就数量而言似乎无关仅要,而事实上,这些氨基酸的合成量已经超过了所估计的需要由饲粮供给的氨基酸维持需要量。 因此,动物真正的氨基酸维持需要量应该是饲粮中回肠真可消化氨基酸需要量与肠道微生物合成量的总和。 对20 kg的猪而言,维持需要的氨基酸量应为1.4 g/d,而不是饲粮提供的304 mg/d。 这说明肠道微生物对动物氨基酸的需要量有净贡献。
同样,饲粮中氨基酸(特别是赖氨酸)的生物学利用率也是一个考虑的重点。 饲料原料的加工、贮存,以及饲料生产过程都会对氨基酸的生物利用率造成负面影响。 很多人错误的认为经化学修饰的氨基酸(chemically altered aminoacids)能在肠道中被消化吸收,只是在动物体内并不能用于合成蛋白质。 事实上,许多经化学修饰的氨基酸不仅不被用于合成蛋白质,而且在到达小肠回肠末端前也不被吸收。 因而,准确预测饲料原料中生物可利用氨基酸含量的一个现实问题就是化学分析方法问题。 在氨基酸的分析过程中,蛋白质的水解是在强酸性体系中进行的,因此,那些已经被化学修饰的氨基酸(不可利用氨基酸)可以转化为原来的活性形式,并被误认为是可利用氨基酸。 从而导致饲料氨基酸含量和氨基酸消化率估计的误差。 现在已经有化学检测方法可以检测饲料中未经化学修饰的赖氨酸(活性赖氨酸)含量。 可是,有证据表明,在动物肠道消化的过程中,并非所有的存在于热损害蛋白质中的活性赖氨酸都被消化吸收。 所以,通过体内法测定饲料中回肠可消化的未经化学修饰的氨基酸,即生物可利用氨基酸才是预测氨基酸消化率的可靠方法(Moughan,2005)。
至今,绝大多数用于预测肠道氨基酸吸收利用的动物生长模型都是建立在试验测定的氨基酸消化率基础之上的。 毫无疑问,这种方法给出的只能是氨基酸吸收利用的近似值,它并不能反应氨基酸吸收利用的动力学过程。 Bastianelli等(1996)建立了数学模型来描述养分消化和吸收过程的机理。 在这个模型中,考虑了肠道食糜不断变化的流动速率,并为进一步引入影响氨基酸消化率的其它因素(包括饲粮和动物)提供了可能。 因果模型可以预测氨基酸消化吸收的动力学过程,从而可以改善预测动物生长的准确性。
4.2. 氨基酸的必然分解代谢(inevitable amino acidcatabolism)
氨基酸的必然分解代谢是指在动物的生长过程中,与能量需要完全无关,仅由于细胞内存在的活性分解代谢酶系引起的饲粮第一限制性氨基酸的分解代谢。 通常认为,当饲粮中除第一限制性氨基酸外的其它养分和非蛋白质源能量均超过动物达到最大蛋白质合成速率的需要时,被吸收的第一限制性氨基酸的利用率应该接近100%。 然而,事实并非如此。
Moughan等(2008)报道,用第一限制性氨基酸为赖氨酸、组成明确的高质量蛋白质饲喂动物,测定赖氨酸在整个机体中的沉积量,并用维持损失的赖氨酸量进行校正。 这一间接测定的结果表明,即使在非蛋白源能量(碳水化合物和脂肪)及其它必需养分充足的情况下,摄入的赖氨酸中大约有15-20%被氧化分解。 近年类似的研究(Mohn等,2000; deLange等,2001)也表明,吸收的赖氨酸和苏氨酸中,其利用率分别为75%和73%。 生长猪回肠真可消化赖氨酸的损耗率为17.4%(Mohn等,2004),这一结果与直接测定的回肠真可消化赖氨酸氧化损失率(13.4%)相当。 色氨酸似乎是利用率最低的一个氨基酸。 吸收的色氨酸中,超过50%没有用于蛋白质合成。 Heger等(2002)的研究也表明生长猪对吸收的色氨酸的利用率也很低(66%),但是赖氨酸的利用率却高达91%。 这一领域的研究同样表明,当必需氨基酸摄入量在维持需要量和约90%最大蛋白质沉积需要量间变化时,每种必需氨基酸的利用率恒定不变(Baker,2005; Heger等,2002,2003),但是不同氨基酸的利用效率却大不相同(Heger等,2002,2003)。
吸收的第一限制性氨基酸为何会氧化损失? 其中一种解释就是基于“氨基酸的必然分解代谢”提出的。 该理论假设,在某种程度上细胞的分解代谢活动是不加选择的,并且氨基酸(即使是蛋白质合成的第一限制性氨基酸)的分解代谢通路并未完全关闭,而是一直存在一定程度的基础分解代谢,并且基础分解代谢的产生是不可避免的。 Stoll等(1999)和LeFloch等(2005)的研究为这一理论提供了支撑数据,表明氨基酸吸收后的分解代谢程度相当大,特别是肠道组织的“首过代谢效应(first-pass metabolism)”。 Stoll等(1998)的一个重要研究表明,28日龄仔猪摄入的赖氨酸中大约有1/3在的肠道组织中代谢。 在“首过代谢效应”利用的赖氨酸中,仅18%用于肠道蛋白质的合成(即这部分赖氨酸被整合到肠粘膜组织中)。 由此推断,有相当部分的赖氨酸由于“首过代谢效应”被分解代谢掉,并且肠上皮细胞可能是赖氨酸及其它必需氨基酸的分解代谢的一个重要场所。
4.3.氨基酸的优先分解代谢(preferential amino acidcatabolism)
氨基酸优先分解代谢过程是阐释饲粮蛋白质-能量互作效应的核心。 蛋白质合成及其它代谢过程需要营养物质分解代谢所产生的ATP提供能量,与此同时,氨基酸也是一个潜在的提供ATP的能量源。 因此,氨基酸既是蛋白质合成的原件,也是潜在的供能物质。 与“氨基酸的必然分解代谢”不同的是,“氨基酸的优先分解代谢”指当非蛋白质原料提供的ATP不能满足动物的需要时,氨基酸则会作为产生ATP的能源物质进入分解代谢过程。
在建立氨基酸优先分解代谢模型时,有三个重要的方面需要考虑。 第一,必需能够预测机体可利用能量的生成速率。 第二,必需对机体每日能量消耗相关的代谢过程进行描述。 第三,为了能够在饲粮能量供给不足时预测氨基酸优先分解代谢的程度,必需提出一些规定或假设,从而可以计算出机体每日蛋白质和脂肪沉积量,也就是说需要通过试验建立一定的数学关系,以便直接预测在能量不足的情况下体蛋白质的沉积速率(Moughan,2008)。
当模型的建立是根据因果关系,而不是以“剂量-效应”试验为基础时,模型将更具有学术性和预测的普适性。 为此,早期的研究者开始寻求建立在因果关系基础上的一些“规则”,以此来描述氨基酸优先分解代谢过程。 动物在生理上对体脂肪含量有一最低需要量(Moughan,2008),这是饲粮蛋白和非蛋白源能量间相互作用的生物学基础。 生长动物必需沉积的最小脂肪含量可能受到诸如品种、性别和年龄的影响。 如果按照“经典”的方法来进行能量分配,则:
那么,根据最低体脂肪含量,动物应该有一个最低体脂肪沉积量; 而剩余的能量却不足以满足最低脂肪沉积所需的能量需求。 此时,氨基酸的脱氨基作用被激活,进一步被分解供能。 这一理论与实际观察的结果一致,即当动物已经贮备了过多的脂肪后,便不会以牺牲体蛋白质(重要器官和骨骼结构必需的)沉积来沉积脂肪。 因而对于体脂肪含量很高的动物而言,在生长过程中允许长期的体脂肪零沉积或负沉积。
5. 结论
建立动物生长的数学模型已经成为动物科学研究的一个主流。 很多学者为描述复杂的生物系统,提出很多理论和模型。 随着对氨基酸消化吸收及代谢过程认识的不断深入,这些理论和模型也在不断改进,并在实践中得到证实。 与此同时,试验研究已经更多的侧重于检验基于因果关系的理论假设,使得试验结果更具有普适性,这为进一步建立更为有效和适用的数学模型奠定了基础。 此外,通过数学模型可以设计出最优收益的饲养制度,因而在商业化的动物生产过程中同样起着重要的作用。
参考文献:略









