春秋战国时期的《墨子·公输》中记载“输攻墨守”法则,比喻只要有一种攻击的战术,就会有防守的方法。同样,在自然界中“输攻墨守”广泛存在,尤其是在宿主与病原菌的“军备竞赛”中表现尤为明显。尽管,众多的科学家明确提出了“输攻墨守”的现象,然而其背后的遗传基础和分子机制任不清楚。
近日,中国农业科学院蔬菜花卉研究所张友军研究员团队在The Innovation(IF=33.2)杂志发表了题为“A midgut transcriptional regulatory loop favors an insect host to withstand a bacterial pathogen”的研究论文。该研究报道了一个中肠转录调控环介导小菜蛾对抗Bt病原菌入侵的新机制。

昆虫是地球上种类最多、数量最大、分布最广的动物类群,但是多数昆虫是农业害虫,给农业生产安全造成威胁。目前,化学杀虫剂是田间防治害虫的主要手段,而大量化学杀虫剂的使用在提高经济效益的同时不可避免导致环境污染和物种多样性破坏的问题。苏云金芽胞杆菌(Bacillus thuringiensis, Bt)是土壤中常见的一类革兰氏阳性细菌,它能产生多种杀虫蛋白,而且基于Bt杀虫蛋白研发的生物杀虫剂和Bt抗虫作物能高效特异的杀死不同农业害虫而对人畜环境安全无害。然而,Bt生物技术产品的大量使用导致害虫抗药性进化的严重问题。据报道,目前全世界至少已经有13种重大农业害虫在田间对Bt生物杀虫剂或Bt抗虫作物产生了抗药性。其中,小菜蛾是第一个被报道在田间对Bt生物杀虫剂进化产生高抗性的农业害虫,是研究害虫Bt抗性机制的理想材料。之前的研究中,张友军团队发现昆虫蜕皮激素(20-hydroxyecdysone, 20E)含量升高可以激活小菜蛾丝裂原活化蛋白激酶(MAPK)信号途径反式调控多个中肠Bt受体和非受体同源基因差异表达,使小菜蛾对Bt Cry1Ac杀虫蛋白进化产生高抗性。然而,导致小菜蛾体内20E含量升高的调控机制尚不清楚。
该研究通过miRNA转录组测序共获得7个在小菜蛾Bt敏感和抗性种群中差异表达的已知miRNAs,利用RT-qPCR分析和功能验证实验鉴定出在小菜蛾Bt抗性种群中肠细胞中显著高表达的功能miR-8545(图1)。
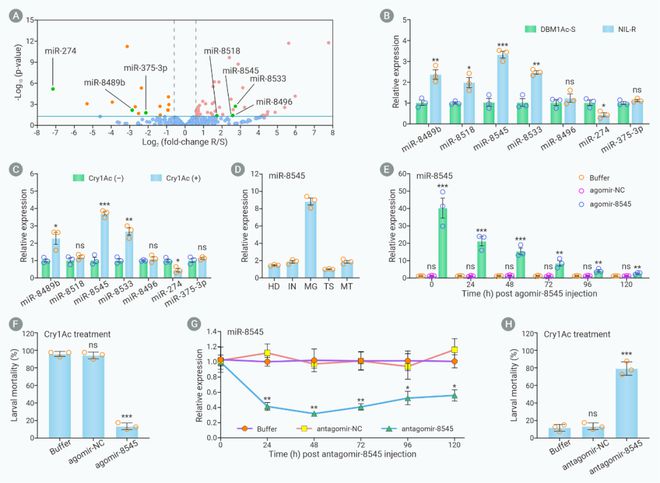
图1. miR-8545表达量增加介导小菜Bt抗性
为了进一步探索小菜蛾中肠细胞中miR-8545通过哪个靶基因介导Bt Cry1Ac杀虫蛋白抗性,作者利用5个miRNA靶基因预测软件共获得7个与miR-8545互作的候选靶基因。RNA免疫沉淀(RNA immunoprecipitation, RIP)、Pull-down和原位荧光杂交技术(Fluorescence in situ hybridization, FISH)实验发现仅有葡萄糖脱氢酶(Glucose dehydrogenase, GLD)基因能与miR-8545直接互作。而且PxGLD基因在小菜蛾Bt抗性种群中肠细胞中表达量显著降低(图2)。
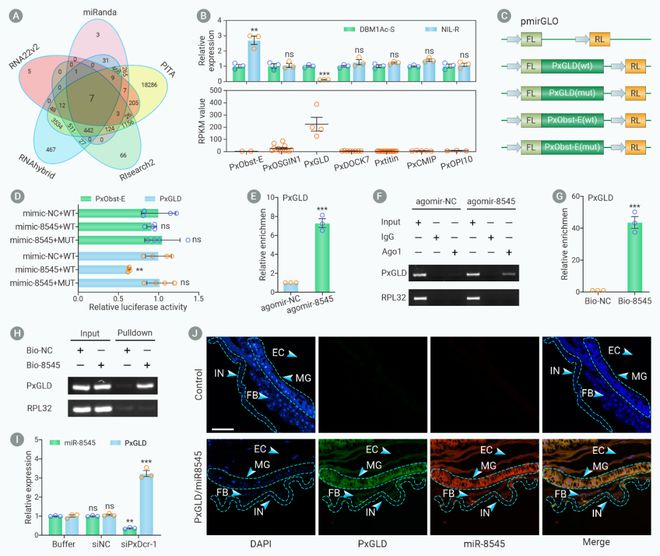
图2. PxGLD基因是miR-8545的直接靶标
为了探索PxGLD基因表达量降低是否参与小菜蛾Bt Cry1Ac杀虫蛋白抗性,利用RNA干扰技术(siRNA)在敏感种群中沉默PxGLD基因后,发现幼虫对Bt Cry1Ac杀虫蛋白的敏感性显著降低。同时,遗传连锁实验发现PxGLD基因表达量降低与小菜蛾Bt Cry1Ac杀虫蛋白抗性紧密连锁。表明PxGLD基因表达量降低参与小菜蛾Bt Cry1Ac杀虫蛋白抗性(图3)。
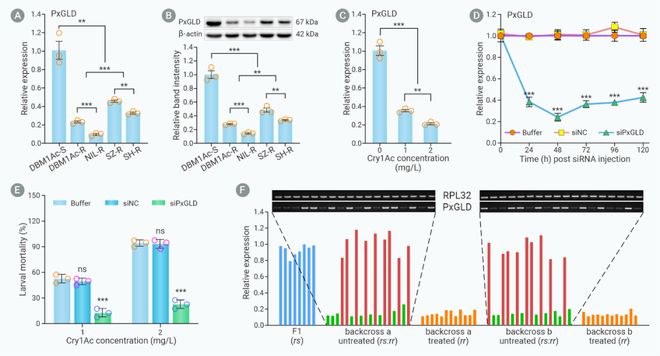
图3. PxGLD基因表达降低与小菜蛾Bt抗性相关
作者接下来探索PxGLD基因表达量降低是否与小菜蛾中肠细胞中20E含量升高相关,结果发现沉默PxGLD基因后20E含量显著升高,同时小菜蛾幼虫注射20E后PxGLD基因的表达量显著增加。UPLC-MS/MS实验检测发现体外表达的PxGLD蛋白能够将蜕皮酮(Ecdysone, E)代谢为3DE (3-dehydroecdysone),将20E代谢为3D20E (3-dehydro-20-hydroxyecdysone)。结果说明,小菜蛾中PxGLD蛋白是一个新的E和20E代谢酶,它的减少导致小菜蛾中肠细胞中20E的含量升高(图4)。
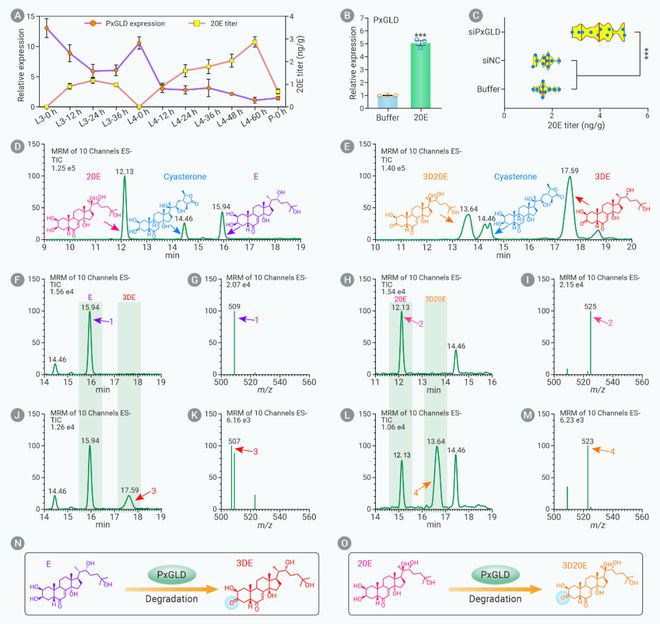
图4. 蜕皮激素代谢酶PxGLD表达量降低导致20E含量增加
为了确认miR-8545是否通过PxGLD/20E模块调控小菜蛾Bt抗性,利用显微注射技术将miR-8545的激动剂注射到小菜蛾敏感种群幼虫体内,结果发现幼虫体内PxGLD的转录和蛋白水平显著降低,20E含量显著增加,生物测定发现小菜蛾幼虫对Bt Cry1Ac杀虫蛋白的敏感性显著降低。相反,在小菜蛾Bt抗性种群幼虫体内注射miR-8545抑制剂后PxGLD的转录和蛋白水平显著增加,20E的含量显著降低,小菜蛾幼虫对Bt Cry1Ac杀虫蛋白的敏感性显著增加。此外,注射PxGLD基因的靶点保护剂后PxGLD的转录和蛋白水平,以及对Bt Cry1Ac杀虫蛋白的敏感性显著增加。结果表明,miR-8545通过PxGLD/20E模块调控小菜蛾对Bt Cry1Ac杀虫蛋白的抗性(图5)。
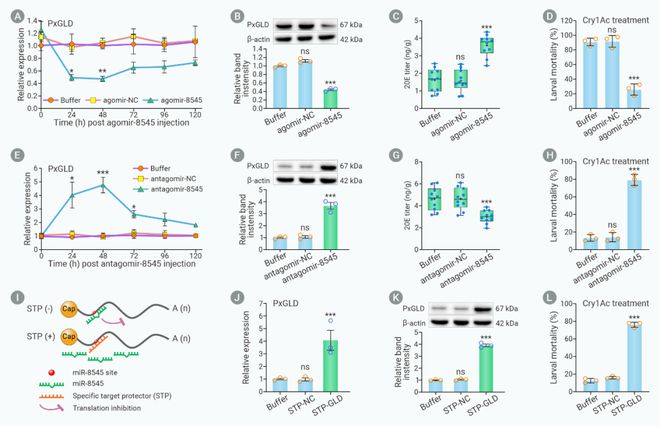
图5. miR-8545通过PxGLD/20E模块介导小菜Bt抗性
为了明确miR-8545/PxGLD/20E模块是否通过MAPK信号级联途径调控小菜蛾Bt抗性,我们通过CRISPR/Cas9技术敲除小菜蛾Bt抗性种群中的miR-8545,结果发现miR-8545敲除后PxMAP4K4基因表达量降低,其它中肠基因的表达量增加,生物测定发现小菜蛾幼虫对Bt Cry1Ac杀虫蛋白的敏感性显著增加。此外,我们发现miR-8545敲除种群中肠组织和蛹体变大。这些结果表明miR-8545/PxGLD/20E模块通过MAPK信号级联途径反式调控中肠受体基因差异表达介导小菜蛾Bt抗性(图6)。
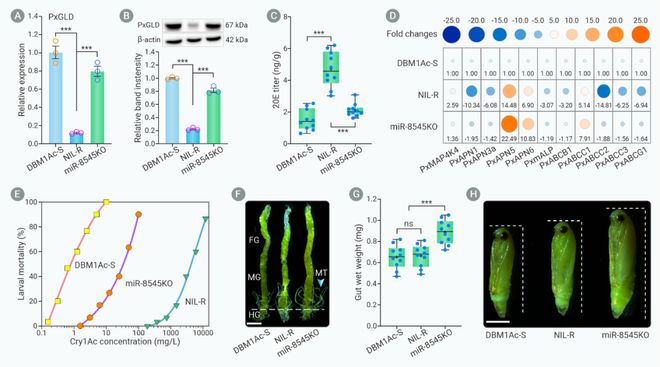
图6. CRISPR/cas9敲除miR-8545后降低小菜蛾对Bt的抗性
为了探索小菜蛾Bt抗性种群中miR-8545表达量增加的上游调控机制,miR-8545的启动子被克隆和进行候选转录因子预测。双荧光素酶实验、电泳迁移率实验(Electrophoretic mobility shift assay, EMSA)和酵母单杂(Yeast one-hybrid, Y1H)实验确定转录因子PxDfd能够与miR-8545的启动子结合。RT-qPCR分析发现PxDfd的表达量在小菜蛾抗性种群中显著降低,沉默PxDfd后miR-8545成熟体和前体及PxMAP4K4表达量显著增加,中肠受体和非受体基因表达量降低。生物测定发现小菜蛾幼虫对Bt Cry1Ac杀虫蛋白的敏感性显著降低(图7)。
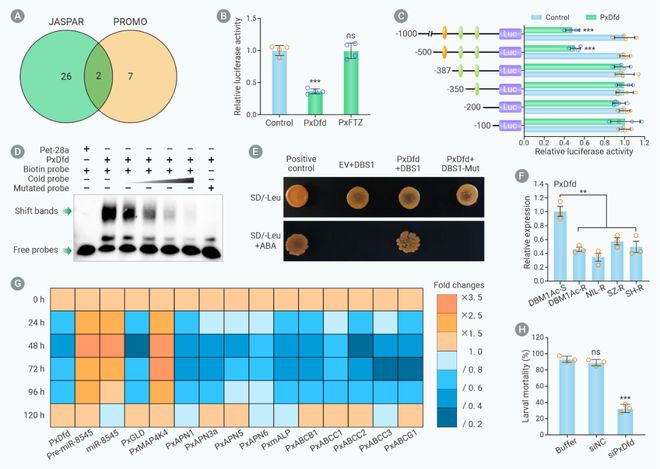
图7. PxDfd通过结合miR-8545启动子介导小菜蛾Bt抗性
作者接下来探索20E是否能通过诱导PxDfd表达量变化介导miR-8545/PxGLD模块的变化。利用显微注射技术将20E注射到小菜蛾幼虫体内,RT-qPCR检测发现20E处理后PxDfd和PxGLD的表达量均显著增加,miR-8545的转录水平显著降低。这些结果表明过量的20E可以通过负反馈机制抑制PxDfd/miR-8545/PxGLD调控轴(图8)。
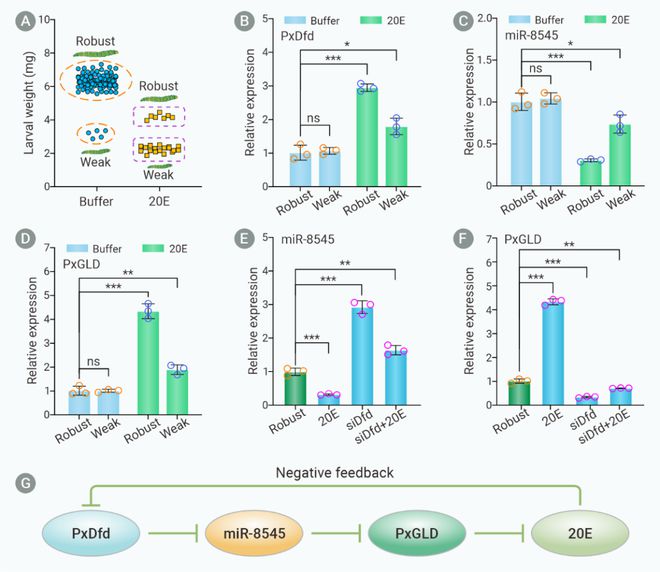
图8. 20E通过负反馈调节PxDfd/miR-8545/PxGLD模块
综上所述,该解析了Bt抗性小菜蛾种群中20E含量升高的分子机制,即PxDfd表达水平的降低导致miR-8545表达量增加,从而抑制PxGLD基因的表达,进而引起昆虫体内20E含量显著升高。随后,增加的20E负反馈调控PxDfd基因的表达,确保小菜蛾中肠细胞中20E的内稳态,使小菜蛾在维持其正常生长发育的前提下进化产生完美的Bt抗性表型。本研究在国际上首次明确了中肠转录调控环介导小菜蛾Bt抗性的分子机制。研究结果对于田间害虫Bt抗性监测预警和综合治理具有重要的理论和实践意义(图9)。
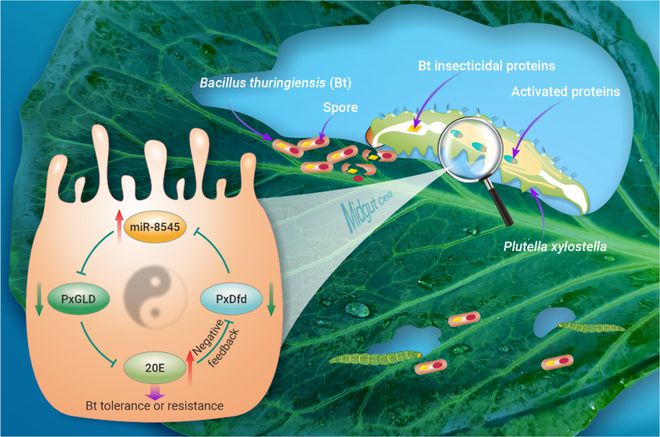
图9. 中肠转录调控环介导小菜蛾Bt抗性的分子机制
中国农业科学院蔬菜花卉研究所的郭兆将研究员、朱流红博士后、程周强博士、董丽娜博士、郭乐博士后(已出站,现为华中农业大学副教授)、白杨博士为本论文共同第一作者,张友军研究员与郭兆将研究员为论文的通讯作者。该研究得到国家自然科学基金创新研究群体项目和中国农业科学院科技创新工程项目等资助。











