在上一篇文章中,我们系统的介绍了多毛纲动物的研究简史,那么这一期文章,我们将带大家去,系统的了解多毛动物的身体构造。
多毛纲动物一般体细长,呈圆柱形或背腹扁平,由数目不等的体节组成,整个虫体通常可分为三个主要部分∶头部、躯干部和尾部。由于虫体各部,尤其是头部和疣足附属物的变化很大,因而多毛纲动物的体型极具多样性,但多种多样的形态都是由基本体型演变而成。
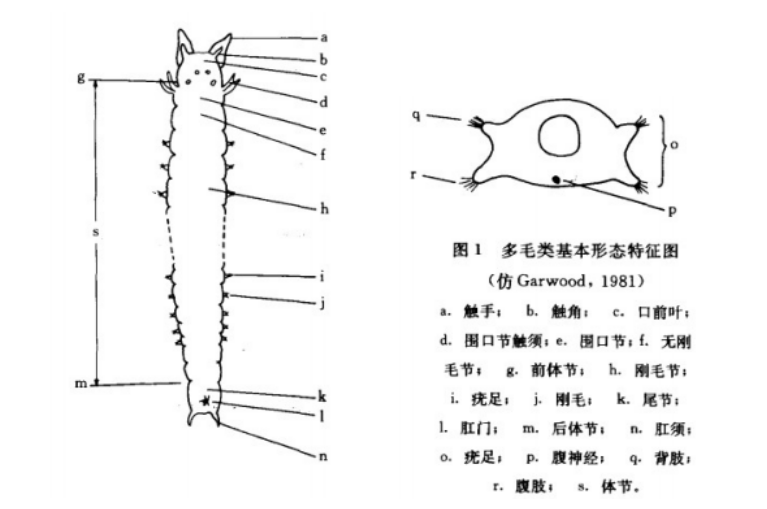
多毛类基本形态特征图(中国动物志无脊椎动物第九卷环节动物门多毛纲叶须虫目)
基本形态的变化包括;口前叶及其附属物的退化;口前叶与围口节的愈合,甚至同前部体节的愈合;体前部捕食附属物的产生,如触须、鳃冠等;躯干部分成不同的区;疣足的退化;刚毛的消失或部分消失;触须、背须和尾须的消失等。
多毛纲动物种间体长变化很大,即使同科不同种间的差异也很悬殊。如,矶沙蚕科Eunicidae 的大矶沙蚕(Eunice aphroditois)体长可达1—3m;而毛轮虫(Ophryotroch puerilis)短于10mm,但该种多数的体长在 10—200mm 之间。

大矶沙蚕(Eunice aphroditois)毛轮虫物种所在的属的物种
底栖生活的叶须虫亚目平均体长为50—100mm,但是也有较大的,如叶须虫(Phyllodoce laminosa) 超过750mm,那么较小的就比如脆拟须虫(Etenides fragilis) 仅为1—2mm。
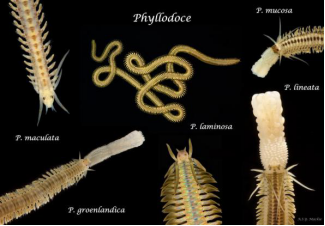
Phyllodoce属物种(中间的为Phyllodoce laminosa),由于脆拟须虫(Etenides fragilis)这个物种比较特殊,因此 并没有找到该种相对应的图呈现给大家
还有,例如浮游生活眼蚕科的细小泳蚕(Plotohelmis tenuis)体长 20一30mm,大型的美丽明蚕(Vanadis formosa) 超过300mm,浮蚕科的最大体长为135mm(Tebble,1962),一般长度为 20—40mm。这样的例子还有很多,在这里我就不一一列举出来了。
体节(segment,somite,metamere)
所有多毛纲动物虫体都有分节现象,沿虫体纵轴,由许多相似的段落或部分组成,而每一段落或部分被称为体节。每个体节两侧,具疣足和背腹须,疣足上又具刚毛。这种体外部的分节和主要器官在各体节内重复排列的现象,被称为分节现象(metamerism, segmentation)。
分节又分为原始分节,次生分节和同律分节,异律分节这两种分节模式。外部与内部体节一致的称为原始分节;次生分节并非虫体真正分节,也称假分节,通常包括体背部不完全的横向沟纹,一般每个体节有 2-3个这种沟纹,也有更多的,在叶须虫科中次生分节常与纤毛带的存在和背部不同的色素相联系。
同律分节(hemonomaus metamerism)
如果所有的体节或多或少有相同的结构,称为同律分节。
我们可以从“同律分节”的字面意义去理解它,“同律”就是相同的规律,“同律分节”便是以相同的规律产生的结构,它们结构表现上近乎一致,比如内部的器官大小,排列等。
同律分节被认为是原始,简单的,是多毛类动物分节的模式。同律分节的多毛类动物所有疣足都具有相同的功能,主要为运动和呼吸功能。最完全和典型的同律分节便出现于叶须虫科中,但真正完全的同律分节是不存在的,叶须虫科前部围口节同虫体其它体节仍有明显差异,其触须比背须更长,后部体节常不发达。
异律分节(heteronomous segmentation)
如果虫体不同部位的体节形态不同,被称为异律分节。
同样,我们也可以从字面意思上来理解异律分节,“异律”就是不相同速率,“异律分节”便是不相同速率的细胞分化,产生的结构也有所不同,比如昆虫,头胸腹等分区。
异律分节在形成的开始阶段,与不同体节的功能划分相联系。在鳞沙蚕科中,一些体节覆盖有背鳞,另一些体节具长触须,这是起补充运动和呼吸功能的作用。还有一些体节有防止阻塞表皮的功能,另一些则是起到了感觉和呼吸功能。
在多毛纲中,矶沙蚕科物种具有律分节到明显异律分节的转变。而只有底栖叶须虫保持了原始的同律分节,这也反映了其原始性。异律分节现象在管栖多毛类中更为显著,例如磷沙蚕科的躯干部可分为前、中、后三区,栖居于复杂构造的U形管中,成虫离开管便无法生存。总之,异律分节发育在不同科中有不同方法,由各种虫体栖息的特殊条件和相适应的生活方式所决定,但异律分节是说明发育的高水平。体节的相异性是由于功能多样性增加,导致结构更多样化,因而外部结构也更趋复杂。
前面,我们说过,多毛纲的虫体通常可分为三个主要部分∶头部、躯干部和尾部。
头部:
头部由口前叶和围口节组成。口前叶由担轮幼虫的前半球发育而成,相对其它体节独立发展,在不同种中口前叶的差异很大。一般说来游走多毛类口前叶较发达,多具有较发达的触角、触手、眼和项器等附属物。而营固着滤食型的口前叶多退化或与围口节和前几个体节愈合,附属物也大都特化为滤食和具呼吸作用的鳃冠(触手冠)。围口节常由1—3 个围口体节组成,有的科围口节由 4个体节组成。围口节腹面具口,两侧多具围口触须。在叶须虫科围口节有的分离,有的愈合,有的保留触须和正常疣足的腹须和刚毛,因此叶须虫科的分类,常用不同的表达式来描述。
口前叶:
口前叶(prostomium) 位于虫体的最前方,为一背腹扁平的多边形或卵圆形的肉质叶。其背面常具1对眼(eye)。
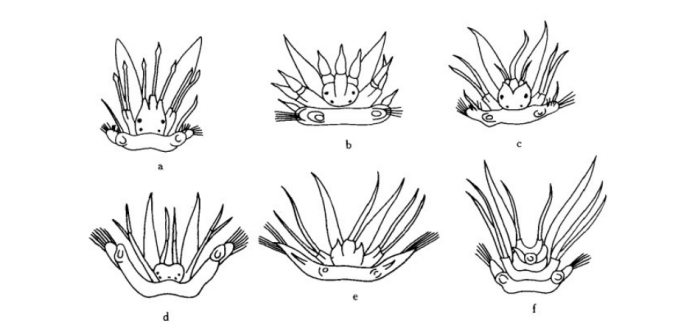
多鳞虫科口前叶的不同类型
眼不仅是多毛纲动物的视觉奇观,同时眼的数目、位置和结构在不同种类中变化很大。例如沙蚕科具4个眼;裂虫科为4一6 个眼;叶须虫科的眼位于表皮下,如同一气泡,其壁由长形的镜状细胞所组成,这些细胞具深色素并形成视网膜,中央具稍可收缩的晶状体,被短的视神经末梢包围,并同中脑相连。眼蚕科的眼很发达且复杂,一般为—对,几平占据了整个头部,眼同肌肉相连,眼轴可改变方向,不仅可看出光线强弱和方向,而且能观察到物体轮廓,Dales (1955)认为眼蚕科主要通过光线来捕食。浮蚕科的眼非常小,同前两科的结构不同,无水晶体;盲蚕的眼完全退化。深海生活和挖掘海底生活的多毛类常无眼。

盲蚕
除了无眼之外,多毛纲有的科则有大量的眼点,如缨鳃虫科中有的种在鳃丝上有大量的眼点;海蛹科的多眼虫(Polyophthalmus pictus)在虫体两侧具12-14 对侧眼。眼的结构简单,仅由感觉细胞和色素细胞组成,甚至只有一感觉细胞。
项器
与口前叶相连的是项器(organum nuchale)又称嗅觉器,通常位于口前叶背面凹陷处。沙蚕科(Nereididae)的项器位于眼后表皮细胞下陷横裂处,内具纤毛,起化学感受器的作用。叶须虫科(Phyllodocida)项器较复杂,除了位于口前叶心形缺刻处的颈部乳突外,还有两个略靠前的内陷侧乳突;背叶虫属(Notophyllum)的项器结构更复杂,数目多达7对。盲蚕科(Typhloscolecidae)项器一对,尤为发达。总之,项器在多毛纲动物普遍存在,是一种化学感受器,具有嗅觉功能,可接受外来的刺激。
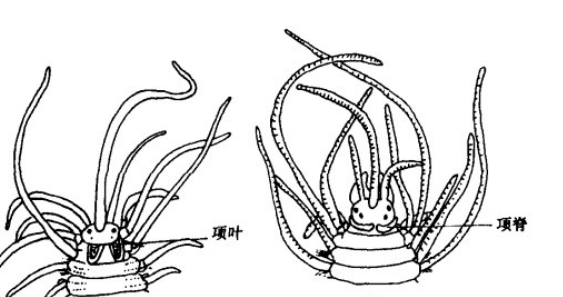
濑户自裂虫(Autolyus setoensis)和叶须真裂虫 (Eusyllis habei)的项器
围口节:
围口节(peristomium)为口前叶后的一个环形节,腹面具横长的口。触须(tentacular cirri) 位于围口节上,在发生上,是与围口节一起由担轮幼虫的前纤毛轮演化而来。触须是特化的疣足背、腹须或须状叶,具很多感觉细胞,可接受外来的刺激。因此具有触角功能。不同科触须数目不同。在沙蚕科触须一般恒定,为3一4 对。叶须虫科触须2—3 对,有的为叶须状。眼蚕科为3—-5 对。浮蚕科触须较特殊,仅2 对,前触须有的短小,位于第1围口节腹面;后触须具内足刺,特别细长,位于第2 围口节,透明浮蚕Tomopteris krampi 的后触须几乎与虫体等长。
触须和触角:
触须触角(palps)与触手(antenna)∶两者在各科命名上及不同多毛类研究者的文献中存在混乱现象,在实际鉴定中往往把两者混淆,尤其初学者,常难于区别,实际上两者有明显不同。头触手由上皮细胞突起形成,具单一神经,受脑控制,位于口前叶的背面或前缘,主要有触觉和化学感受器功能。触手的数目在游走多毛类中较稳定;沙蚕科 2个;海女虫科 2—3个;裂虫科和鳞沙蚕科 3个;吻沙蚕科和齿吻沙蚕科4个;沙蚕科则可能1个,3个或5个;叶须虫科为4—5个;眼蚕科5个;特须虫科和盘首蚕科为4个;须蚕科1对;无指蚕科和浮蚕科无触手。触角与触手的区别在于体腔进入触角,触角分别由脑和围咽神经环的两条神经支配,它比触手更为敏感,其主要功能是感觉和捕食作用。触角在不同科间有变化∶海稚虫科常有成对具沟的粗触角;浮蚕科具1对宽大的触角,是分类的重要依据;沙蚕科的触角具基节和端节;而叶须虫科、眼蚕科、盘首蚕科、盲蚕科和特须虫科的触角消失。
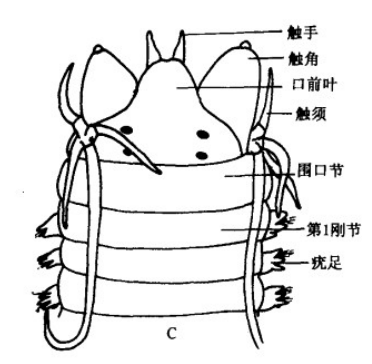
双齿围沙蚕 Perinereis aibuhitensis的头部躯干部:
躯干(trunk)及其附属物位于头部之后,尾部前面。在发生上,躯干部是由担轮幼虫的后半球发育而来,但它不包括尾部,更确切的是指包括在尾部前的生长区及随发育形成的体节。体节可能彼此相似或不同。由于前后体节结构的不同,导致躯干部出现分区现象,形成体前部(胸区)和体后部(腹区)甚至更多区。躯干部最主要的附属物是体节两侧具疣足,疣足上具刚毛。
疣足:
疣足(parapodium) 位于多毛类的体节上,由体壁向体侧伸出的突起物,具有运动、呼吸、排泄、生殖和保护等功能,但主要是行运动功能。在胚胎发育过程,疣足是在幼虫发育进入疣足幼虫期以后才逐渐分化形成的。疣足发育的最终结构取决于多毛类的栖息环境特征以及相适应的生活方式。
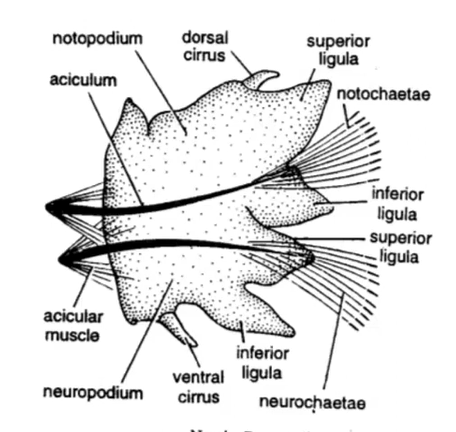
疣足的详细结构在多毛类分类各阶元中变化较大。依结构不同,分双叶型、单叶型和亚双叶型三类。典型疣足为双叶型,具背叶和腹叶、背刚毛和腹刚毛、背须和腹须、背足刺和腹足刺。单叶型疣足无背叶和背刚毛,亚双叶型疣足背叶退化仅余背刚毛或背足刺。

叶须虫目的疣足
多毛动物疣足的背叶和背须变化在超科和科的分类阶元中较为重要,在鳞沙蚕亚目中有的背须特化为鳞片,从而具有保护功能;叶须虫科的背须呈叶片状,成为游泳桨,便具有了运动功能;沙蠋(音同“竹”)科(Arenicolidae)虫体的背须上布满细血管,具有呼吸功能;而眼蚕科水蚕属的雌性虫体前两个疣足的背、腹须特化成受精囊,具有生殖功能;盲蚕科疣足叶退化,只留下1一3 根短小的简单型刚毛,背、腹须膨大成宽板,使身体在水中保持垂直;浮蚕科的疣足无刚毛和足刺,只有背腹足干和足膜,更适应浮游生活。原环虫纲(Archiannelides)的 疣足退化为小的乳突。总之,多毛纲的疣足结构千变万化,成为多毛纲分类学上的重要鉴别之一。
刚毛:
在上期,我们曾提到多毛纲,因其具刚毛而得名,那么刚毛是什么?
刚毛(seta,chaeta,bristle)是位于多毛动物疣足叶外部或内部的几丁质刺毛,由刚毛囊(setal sac)的毛原细胞(formative cell)分泌形成。具辅助运动、保护、生殖或捕食的功能。刚毛有两类,位于疣足内的通常较粗称足刺(aciculum)或称内刚毛。内刚毛(足刺)是疣足的支撑轴,它通过特殊的肌肉可轻微的伸缩,从而控制外部刚毛的活动,具有辅助运动功能。内刚毛通常为黑色,在每一刚叶内部有1至2根,眼蚕科泳蚕属等的足刺常伸出刚叶的外部,是分类的重要依据。
足刺(aciculum):疣足内部较粗且色较深的刚毛称为足刺,同时也被称为内刚毛(inner seta)。足刺具足刺肌,并支撑着疣足和其他刚毛。
由疣足外伸成束的刚毛称外刚毛,通常简称刚毛。外刚毛在形态上变化较大,根据刚毛的结构可将其归纳为两种主要类型,刚毛不分节的为简单型,分解的则为复型。除了简单型和复型之外,刚毛还有一个次要类型即伪复型刚毛,伪复型刚毛分节不明显,并且也较为少见。
那么,这些刚毛又有哪些具体表现呢?
简单型刚毛(simple chaetae):
简单型刚毛就是不分节或不具关节的刚毛,有毛状、梳状、亚足刺状、刺状、桨状、镰刀状、匕首状、钩状、鸟喙状、齿片状等。如日本刺沙蚕(Neanthes japonica)体后部腹刚叶上的简单刚毛、双管阔沙蚕(Platymereis bicanaliculata)体中部和后部背刚叶的鸟嘴状刚毛、美丽阔沙蚕(Platymereis pulchella)体后部的具肋刚毛等。
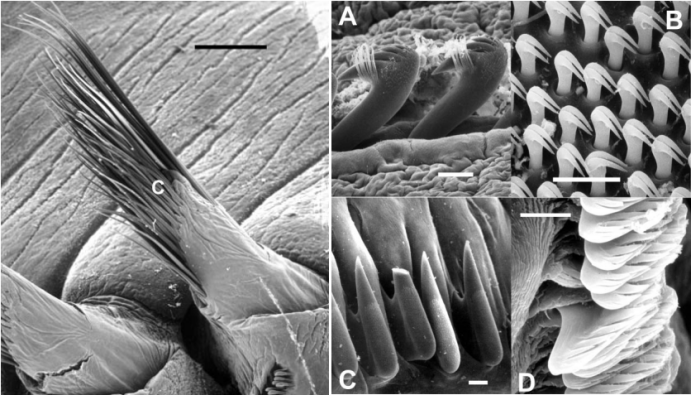
左C为毛状刚毛,右ABCD则为勾状刚毛
复型刚毛(compound chaetae ) :
具分节或具关节(joint)的刚毛。其基部为柄(proxi-mal shaft),前端称端片(distal blade)。依端片形状(刺状、镰刀形、桨状)和柄前端两齿等大(等齿)或不等大(异齿)的组合,复型刚毛又可分为;异齿刺状刚毛(heterogomph spinigerous seta),等齿刺状刚毛(homogomph spinigerous seta),异齿镰刀形刚毛(heterogomph flagerous seta),等齿镰刀形刚毛(ho-mogomph flagerous seta)和桨状刚毛(paddle seta)
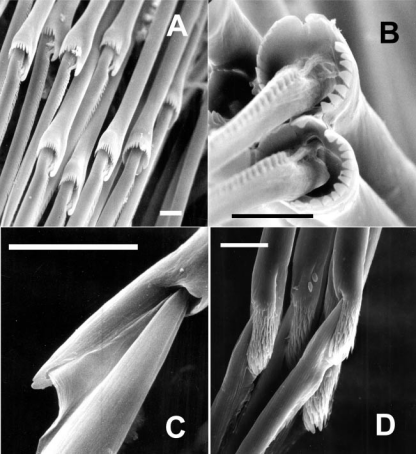
各种复型刚毛
伪复型刚毛(pseudo-compound seta):
又称半齿刚毛(hemigomph seta)。刚毛端片与柄部大部愈合,但有分界线,为介于简单刚毛和复型刚毛间的一类刚毛。
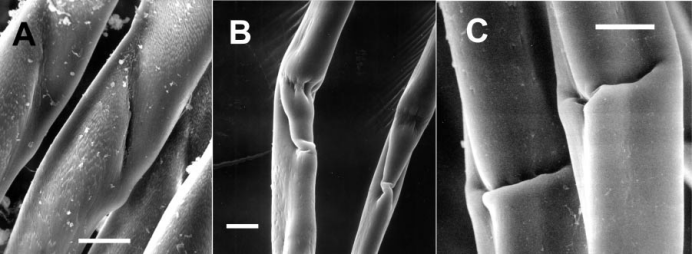
伪复型刚毛
在发生上,所有多毛类的刚毛都在刚毛囊中发育,刚毛囊则是表皮的内陷。单叶型疣足只有一个刚毛囊。刚毛囊内部具许多囊状细胞并有膜相连,囊状细胞的分泌物形成刚毛基底细胞,每一个基底细胞常形成一根刚毛(见VmakoB,1972)。刚毛囊的基部是产生新基底细胞的发生区,保持了新刚毛替代消失的刚毛。基底细胞有一大的细胞核和一滤泡。根据 Bobin(1944)的研究基底细胞还有一特殊的具毛的生长体颗粒,在刚毛生长中具毛的生长体颗粒增多,并伸出细小的纤维进入细胞中,以后形成新刚毛。复型刚毛也同样在刚毛囊中发育,不过刚毛远端部分先形成,柄部后形成,柄部完全形成后才穿透表皮。刚毛束是新刚毛的发生和发育区,它保证了新刚毛替代消失的刚毛。
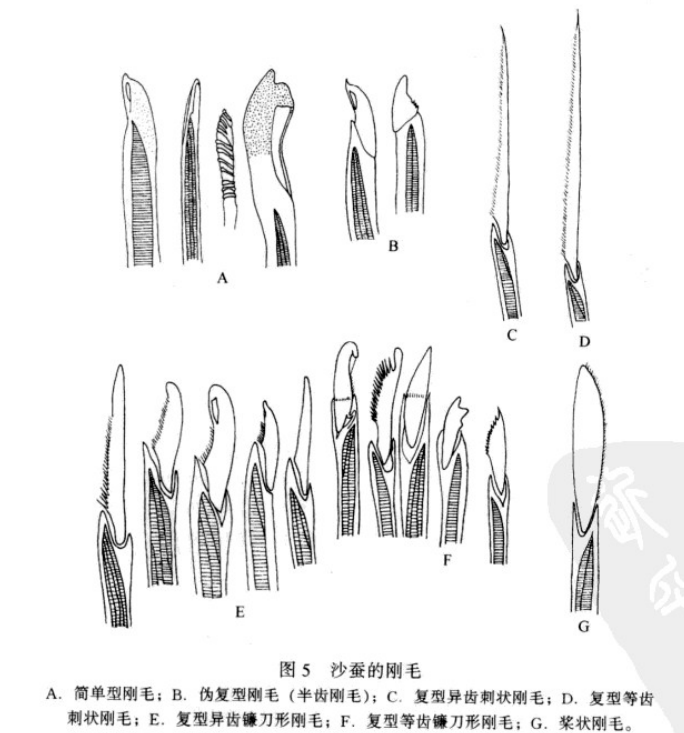
沙蚕的各种刚毛(中国动物志无脊椎动物第九卷环节动物门多毛纲沙蚕目)
总之,刚毛的各种性状是多毛类分类学上重要鉴征之一(详见各科的描述),深入的研究刚毛仍为多数分类学家所关注。观察乃至粗略地鉴定多毛类标本都少不了使用显微镜仔细地研究刚毛,若能使用扫描电镜精细研究刚毛形状,将有助于正确的进行分类。

显微镜下的各种刚毛
尾部:
尾部(pygidium)是位于虫体最后的一节或数节,又称肛部。在发生上,尾部是由担轮幼虫后半球演化而来。尾部上常有数目不等的尾须或其它精细结构,对于盲蚕科和裂虫科、海稚虫科、海蛹科、沙蝎科、缩头虫科(Maldanidae)、笔帽虫科、缨鳃虫科(Sabellaridae)、等物种来说是重要的鉴征,若是失去了尾部便会造成无法鉴定种或属的悲剧发生。
笔帽虫科的尾部由 5—6个体节组成特殊的附属物;缨鳃虫科的尾部为香肠状,尾部朝向栖管的开口处,这种结构可以让排遗物直接排入海中;盲蚕科的尾部有两个大叶状肛须,很像鱼的尾鳍,无疑是对浮游生活的高度适应:而叶须虫亚目尾部变化则较大,在此不便于描述,仅提供图片给大家进行参考。
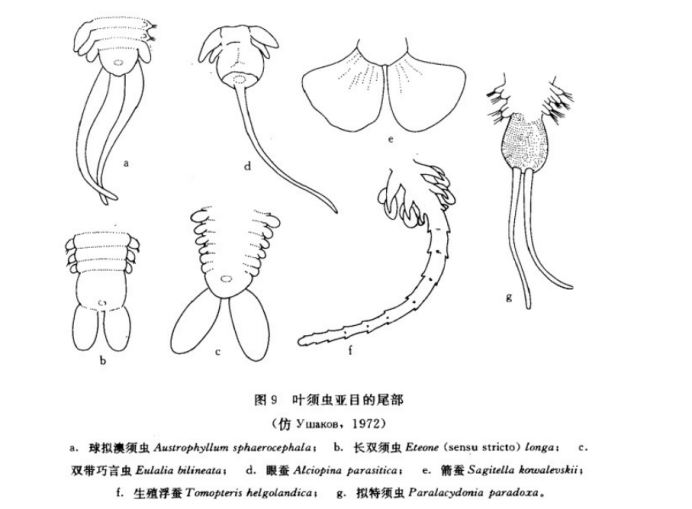
图源中国动物志无脊椎动物第九卷环节动物门多毛纲叶须虫目文献引用:
中国动物志_无脊椎动物_第九卷_环节动物门_多毛纲(一)_叶须虫目
中国动物志_无脊椎动物_第三十三卷_环节动物门_多毛纲_(二)_沙蚕目
中国动物志无脊椎动物_第五十四卷_环节动物门_多毛纲(三)_缨鳃虫目
Polychaete chaetae: Function, fossils, and phylogeny_Rachel A. Merz1, and Sarah A. Woodiny 2006
Taxonomy at order and family levels of the benthic groups of Polychaeta in the coastal waters of China_Meiling Ge , Qinzeng Xu, Shiliang Fan1 , Zongxing Wang , Xuelei Zhang 2018
The Ophryotrocha labronica group (Annelida:Dorvilleidae)with the description of seven new species_HANNELORE PAXTON&BERTIL AKESSON
Two Species of Fireworms(Annelida:Amphinomidae: Chloeia) from Hong Kong_Zhi Wang, Yanjie Zhang, Yang James Xie, and Jian-Wen Qiu
