题目:Overexpression of SHORT VEGETATIVE PHASE-LIKE (SVL) in Populus delays onset and reduces abundance of flowering in field-grown trees
刊名:horticulture research
作者:Amy L. Klocko , Steven H. Strauss et al
单位:Oregon State University, Corvallis
日期:01 August 2021
01
摘要
转基因和外来种质从种植作物传播到野生或野生物种是公众和监管机构接受基因工程植物的一个难题,特别是对于杨树等风媒树。我们报道了花抑制因子短营养期(SVL)的杨树同系物的过度表达,拟南芥 MADS-box 抑制因子短植物期(SVP)的同源物,在田间的三种基因型中延迟了数年的开花的转基因杨树。SVL的高表达与开花开始延迟和花数量降低相关,并且不会对叶特征、树形或茎体积造成形态学上明显或统计学上显着的影响。过度表达对春季生殖和植物物候的影响是适度的,并且是基因型特异性的。我们的结果表明,使用SVL和相关的花抑制因子可以成为对无性繁殖的短周期木本能源或纸浆作物进行高水平遏制的有用工具。
02
技术路线
果杨(Potri.007G010800)的SVL基因被克隆到过表达载体 pCAPO 中
杨树无性系与遗传转化
田间种植
茎生长和叶片特征、花卉尺寸
基因表达分析
统计分析
03
主要结果
3.1SHORT VEGETATIVE PHASE-LIKE (SVL)的鉴定和过表达
由于在抑制开花起始基因中的作用,我们试图确定与拟南芥SVP基因最接近的毛果杨同源物。SVP和AGL24是StMADS11超进化枝中唯一的拟南芥成员,而毛果杨基因组包含九个成员(图1A)。然而, SVP亚进化枝中只有一个杨树基因组。SVP 直系同源物的编码序列,称为SVL ,是从P. trichocarpa cv.“ Nisqually-1”中克隆的。然后放置在35S的下游具有重复增强子元件和章鱼碱合酶终止子上游的启动子(图1B)。SVL的表达值和P. trichocarpa中最接近的相关 MADS-box 基因在几种组织类型中的表达值显示在附件中。
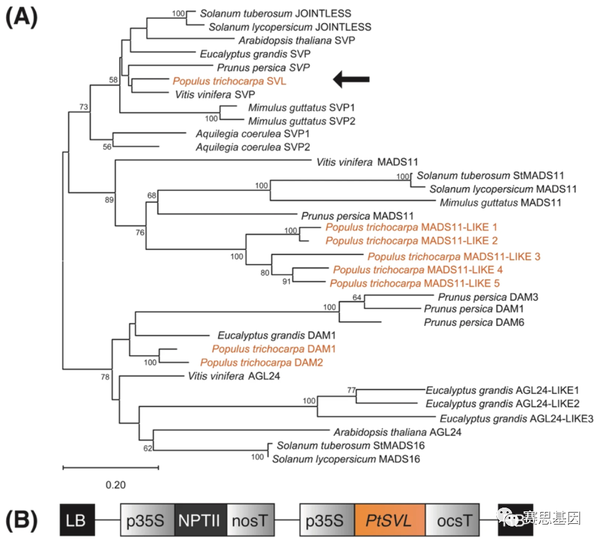
3.2 35S:SVL转基因表达及八年生长田间观察
从 2011 年到 2019 年,所有无性系的实地树木存活率为 95% 。我们从 2014 年开始测量花卉丰度,当时种植园中的大部分树木开始开花。在 2016-2019 年,当许多树木开花普遍且密集时,我们注意到许多35S:SVL树木不开花的例子,但相邻树木花量丰富(图2B-D)。虽然邻近的树木也是转基因的并且包含旨在诱导不育的结构,但这些邻近的树木被确定具有与基于表型评估的非转基因对照相似的花期。虽然我们没有注意到35S的植物外观有任何明显的表型差异,我们观察
到35S:SVL树上冠的次级分枝减少的趋势(图2B-D)。

图 2:田间种植的SVL过表达转基因杨树的代表性图像。
我们在 2018 年春季评估了对照树和转基因树中的SVL基因表达,发现我们的过表达构建体导致所有三个测试克隆的表达增加。在 353 克隆背景中发现了具有最高表达的事件(图3)。在717背景中,我们发现了一些高表达的事件和许多低表达的事件(图3)。在大多数情况下,SVL表达在同一事件的两个分株中相似,但在少数情况下(例如,353:28、717:54),SVL表达在分株之间存在显着差异(图3 ). 在考虑了涉及克隆、构建体和块的混合效应后,我们发现 70% 的基因表达差异发生在事件之间,30% 发生在事件内的分株之间(即,包括分株之间和块之间的变异)。
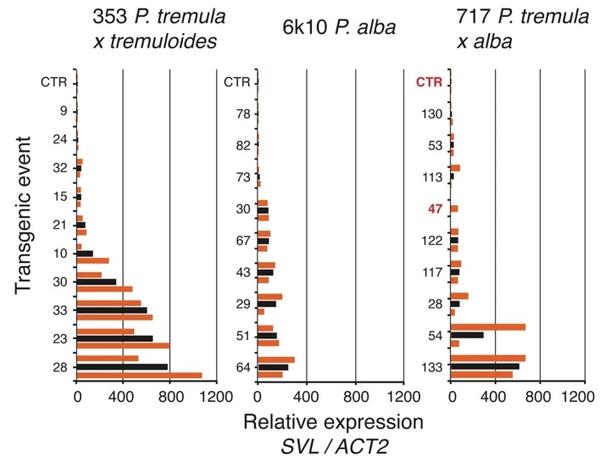
图3:SVL在过表达杨树转基因品系中的相对表达。
3.3 SVL表达与花期延迟和花丰度降低相关
转基因田间种植的环境变化导致树木生长速度的巨大变化,从而导致繁殖的开始。为了控制这一点,我们对茎体积进行了空间调整,如前所述。为了确定SVL对改良繁殖的有用性,我们使用线性回归来测试SVL表达是否与开花的开始或丰度相关。在初步分析显示体积和花卉性状高度相关后,我们在所有回归分析中使用茎体积作为协变量。我们发现SVL表达与开花开始之间存在高度显着的关系(Pr < 0.0023)(图4A和补充表2 )). 我们发现SVL表达与花丰度(树冠中的柳絮覆盖率;参见“方法”部分,图2 )之间存在适度显着的关系(Pr < 0.0219)(图4B)。
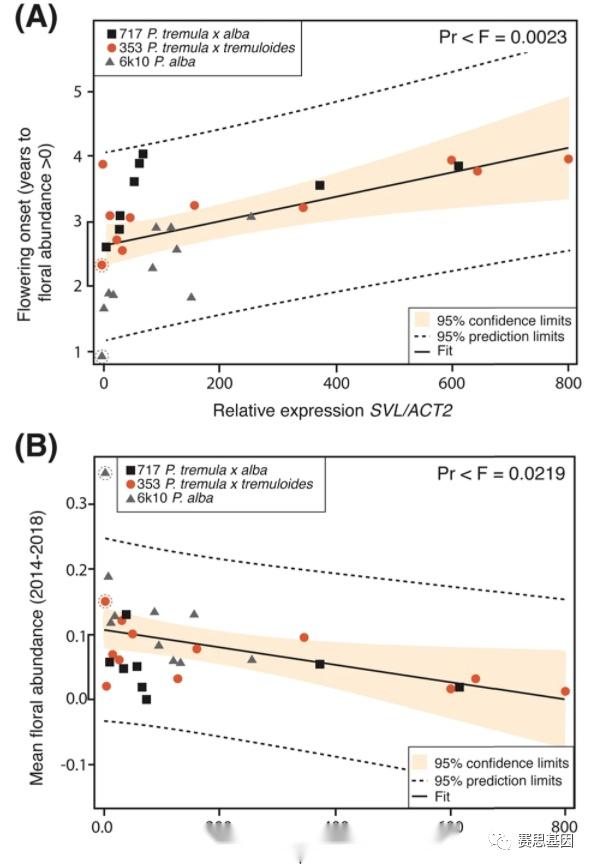
图 4:杨树转基因和对照品系中开花性状与SVL表达的回归分析。
A开花开始(年)与SVL基因表达相对于ACT2持家基因。
B花卉丰度(所有年份的平均花卉丰度)与SVL相关基因表达
基于SVL的分子功能,我们预测SVL的高表达会延迟开花和丰度33。因此,对于每个克隆,我们比较了随时间变化的三个最高和三个最低SVL表达事件的花丰度(图5)。对于每个克隆,低表达事件始终具有更高的估计花丰度(针对每棵树的体积指数的变化进行协变量校正;图5)。35S:SVL转基因的一般花序形态在 353 和 6k10 克隆的强制休眠插条中进行了评估,与对照相比是正常的(补充图8A,B ).
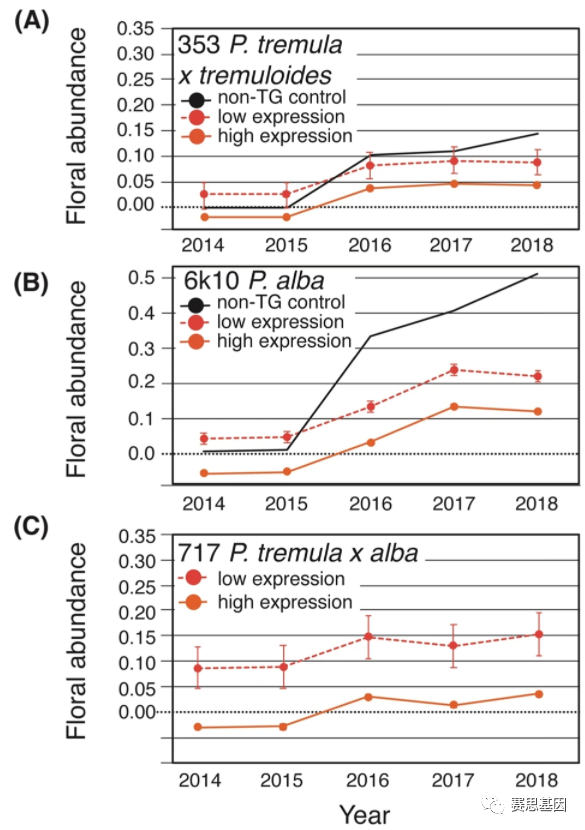
图 5:低表达和高表达35S:SVL系(事件)随时间变化的花丰度。
3.4 SVL过度表达的树木具有正常的营养性状
在八年的生长期,我们收集了茎体积、叶面积和质量、叶柄长度和宽度以及叶绿素含量的数据。为了研究SVL表达对叶片营养特性的影响,我们使用以茎体积作为协变量的相同线性混合效应模型分析了 2018 年的叶片表型。我们的回归分析没有提供强有力的证据证明SVL表达与任何叶片性状之间存在统计学上的显着关系(补充表1;所有 Pr ≥ 0.0975);所采用的统计分析的例子在 Suppl 中针对十个特征进行了介绍。然而,使用没有克隆的模型,对大多数性状的影响在大约 10% 的水平上是显着的(补充表1A)。从叶柄和叶密度比L:W的回归截距可以看出,营养性状在克隆之间差异很大,但在克隆内差异很小。因为体积是我们统计模型中的一个重要协变量,所以我们使用了几种方法来研究SVL表达对茎体积的潜在影响。首先,我们对茎体积与SVL表达进行了回归分析。其次,我们进行了相同的分析,但将克隆作为固定效应包括在内(补充图3B)。在这两种情况下,都没有统计学上显着的关系(P > 0.13)。对于克隆717,体积指数与SVL表达呈负相关的趋势较弱;然而,它显然取决于体积非常低且SVL表达高于平均水平的单个事件(补充图3B;Pr < 0.13)。
3.5 35S:SVL转基因在花和营养芽断裂方面表现出较弱的基因型特异性延迟
有报道称SVL可以调节杨树的营养萌芽。我们通过在 2014 年和 2015 年的 1 个月期间对营养性芽破进行评分(补充图4),通过在整个不育试验(包括其他结构)中定性评分与正常花芽破发时间的偏差,对芽破进行了有限的观察2018 年(补充图5),并通过对 2016 年初春 6K10 转基因植物的详细花芽断裂形态进行评分(补充图6)。在 717 中,但在 353 或 6K10 中,我们观察到 2014 年和 2015 年35S:SVL的营养萌芽延迟转基因相对于对照。对于 2018 年的花芽断裂,在不育试验中,9% 的存活树木被评为异常晚花芽断裂。这包括 9 个35S:SVL转基因分株(试验中 179 棵此类树中的 5%),而效果最强的构建体 PFPG 和 PLF(单独靶向LEAFY或与AGAMOUS结合使用 RNAi 击倒) , 延迟潮红的发生率高出 6 倍;他们的 210 棵树和 188 棵树中有 24% 分别被评分为迟到(补充图5)。35S:SVL转基因是晚期花芽断裂不育试验中 24 种构建体中的第 14 种,远低于显着晚期 PLF 或 PFPG 构建体,并且显示与确定表达的那些树的SVL表达无关(Suppl. Fig. 5 B , C)。基于 2016 年 6K10 35S:SVL事件和对照中柔荑花芽形态的详细评分,我们发现转基因事件中花芽断裂的延迟较弱但在统计学上不显着(补充图6A);此外,在可获得表达信息的分株中,花芽断裂与SVL表达的相关性很差,并且通常比对照树延迟更少。
04
结论
SVL表达的修饰可用于控制杨树的开花,我们的研究表明, SVL过度表达会导致首次开花的强烈延迟,但对物理特征、叶片形态或营养生产力没有显着影响。
05
原文获取
原文链接:
nature.com/articles/s41438-021-00600-4
PDF获取:
scientsgene.com/h-nd-151.html
