麦康森
中国海洋大学
中国工程院院士
导读:蛋白质是水产饲料的主要营养物质,也是决定养殖成本的关键因素。水产动物对蛋白质含量要求较高,一般是畜禽的2~4倍,鱼粉含有高质量的蛋白质和必需氨基酸以及未知生长因子,具有良好的适口性、较高的消化吸收率,是水产饲料中最重要的蛋白源之一。目前,全球三分之二的鱼粉都用于水产饲料,但随着水产养殖业的不断发展,鱼粉等优质蛋白资源不能满足市场需求的情况日益加剧。替代鱼粉,提高蛋白利用率和开发新的蛋白源,是行业面临的核心问题。2021年第一期我们特别邀请中国海洋大学麦康森院士以“水产饲料的蛋白源问题——提高饲料蛋白质利用率新思路”为题,介绍鱼粉蛋白与其他蛋白的组成成分的差异,阐述提高饲料蛋白质利用的理论基础;并在深入探讨影响鱼体蛋白沉积潜力内在的因素下,提出激活水产动物mTOR信号通路和/或消除其抑制因素,为提高饲料蛋白质利用率提供了新思路,供行业同仁学习借鉴。 全文已在《饲料工业》2021年第1期刊出。
蛋白质是生命的物质基础,没有蛋白质就没有生命。鱼粉是水产饲料中最重要的饲料蛋白源之一,国际上超过2/3的鱼粉被用作水产饲料,在中国达60%左右。鱼粉主要用于水产饲料,其次是猪和宠物的饲料,禽类饲料中较少使用。水产饲料配方中严重依赖鱼粉,因此谈到水产饲料蛋白源就一定会涉及鱼粉。根据战略分析报告,随着人口的增长,人们对水产品的消费需求越来越多,预计未来鱼粉产量将会维持在500万吨左右,不可能再继续增加。加之水产养殖业的发展对鱼粉的需求越来越大,因此水产养殖营养学家们一直在寻找替代鱼粉和减少鱼粉使用量的解决方案,即研究如何提高现有鱼粉的利用率或者开发新的蛋白源来替代鱼粉,鱼粉替代已是一个世界性难题。
提高饲料蛋白质利用率的理论基础
鱼粉是目前应用最为广泛的水产饲料蛋白质,与其他蛋白源(植物蛋白、陆生动物蛋白、单细胞蛋白、昆虫蛋白)具有显著的区别。研究表明,以非鱼粉蛋白源为主要蛋白质原料时,即使在氨基酸、能量、脂肪酸、无机盐和维生素等组成方面都与鱼粉为主要蛋白源的饲料尽可能一致,水产动物的生长还是比不上鱼粉饲料,即饲喂鱼粉饲料比饲喂其他蛋白源饲料的水产动物具有更好的生长优势,这表明除了已知的营养成分外,鱼粉中还必然存在一些其他蛋白源没有的未知生长因子。过去,人们往往主要考虑蛋白源之间的差异导致的水产养殖动物生产性能的不同,即考虑影响生产性能的外因较多,而对水产动物本身对不同蛋白源饲料的消化吸收和代谢的内在反应——内因研究较少。因此,同时考虑研究外因与内因对饲料蛋白质利用率的影响具有重要的理论价值和现实意义。
1.1 鱼粉与其他蛋白源的组成差异
在开展鱼粉与其他蛋白源组成的差异研究中,通常根据现有的营养学知识,比较研究其适口性、主要营养成分(蛋白质、氨基酸、脂肪、脂肪酸、矿物质和能量等)、营养拮抗因子和消化率等指标。研究发现,鱼粉中营养成分含量丰富,而传统上认为一些非必需的物质,在替代鱼粉的饲料中却起到非常重要的作用。当然,在非鱼粉蛋白源中,尤其是植物蛋白源中同样存在一些未知的营养拮抗因子。逐步探索研究正反两方面的未知因子的作用机理,对最终解决鱼粉替代的难题至关重要。
1.1.1 牛磺酸
鱼粉中富含牛磺酸,是其他蛋白源中含量较低甚至缺乏的重要活性物质。在不同的饲用蛋白源中,牛磺酸在鱼粉中的含量最高,其次是肉骨粉、酵母,在植物蛋白中含量少。因此,用不同蛋白源设计配方时,非鱼粉蛋白源中牛磺酸就会显著不足。牛磺酸在鱼体内合成能力差,尽管不参与蛋白质的合成,但对于维持鱼体渗透压平衡、抗氧化、抗应激等具有重要作用。此外,牛磺酸对鱼机体神经系统、心血管系统、免疫系统、肝脏、肌肉等也具有非常重要的作用。当用非鱼粉蛋白源饲料饲喂时,由于牛磺酸不足,就会逐渐暴露出牛磺酸缺乏引起的营养问题。进一步研究发现,大多数鱼类自身合成牛磺酸的能力有限。究其原因,自身合成牛磺酸的途径一般需要以蛋氨酸、半胱氨酸为前体,经过半胱亚磺酸脱羧酶等关键酶催化成亚牛磺酸,进而形成牛磺酸。但大多数鱼类上述代谢通路的活力非常弱,远不及大鼠、小鼠等物种。深入探究发现,大多数鱼类中牛磺酸合成的关键酶——半胱亚磺酸脱羧酶的底物结合位点发生突变,牛磺酸合成效率低,尤其是海水鱼更为严重。因此水产饲料中必须含有一定量的牛磺酸,而这个来源最直接的就是饲料成分鱼粉。因此,在非鱼粉蛋白源为主的水产饲料中,必须要考虑补充添加牛磺酸的问题。
1.1.2 羟脯氨酸
羟脯氨酸是构成胶原蛋白的主要成分,高等动物可以由脯氨酸羟化酶催化脯氨酸形成,但鱼类羟脯氨酸的合成能力较弱,需要外源补充。鱼粉同时也是活性物质——羟脯氨酸的优质来源,植物蛋白和其他蛋白源含量很低。当饲料中的鱼粉被植物蛋白替代到一定水平后,鱼体羟脯氨酸的水平会出现明显下降,进而可影响胶原蛋白的合成与生长。因此,要实现鱼粉的有效替代,饲料中额外添加一定量的羟脯氨酸就显得非常重要。实验发现,当饲料中60%的鱼粉被植物蛋白替代后,鱼体的特定生长率显著下降,补充0.6%的羟脯氨酸可显著改善其生长,并提高血浆和肌肉羟脯氨酸含量以及肌肉胶原蛋白含量。此外研究发现,饲料中充足的羟脯氨酸含量对于鱼体背肌硬度、咀嚼度、弹性等品质指标均具有改善作用,这与羟脯氨酸是胶原蛋白的组成成分有关。
1.1.3 维生素D3
众所周知,维生素D3对于水产动物营养具有重要的意义,这与钙、磷的吸收有关。植物来源与鱼粉、鱼油来源的维生素D存在显著差异,植物来源的一般为维生素D2,鱼粉鱼油来源的是维生素D3。研究表明,相对维生素D2,维生素D3对促进钙磷吸收、骨骼矿化、免疫分化更有效。研究发现,维生素D3与脂肪代谢也有显著关联,维生素D3缺乏可显著抑制鱼类生长。维生素D3能够通过诱导过氧化物酶体增殖物激活受体γ-1α(Pgc1α)的表达,促进体脂肪的氧化供能利用,提高蛋白质沉积率。当维生素D3缺乏时,鱼体腹部脂肪不能转化为能量而大量沉积。因此,在鱼粉和鱼油用量不高的水产饲料中,必须要补充添加维生素D3。
1.2 各类抗营养因子
研究发现,与鱼粉相比,豆粕等植物蛋白中存在多达十几种鱼粉没有的抗营养因子。目前已经证实这些抗营养因子可产生毒害作用,如大豆皂甙可引起消化道黏膜上皮组织损伤、微绒毛脱落;棉酚可引起鱼类肝脏纤维化等一系列的问题。因此要实现植物蛋白源替代鱼粉,必须明确这些植物蛋白源中抗营养因子的负面效应和作用机理,以针对性研发出相应的消除或钝化技术。
1.3 体蛋白质沉积潜能差别
鱼粉与其他饲料蛋白源相比,其在蛋白质含量、氨基酸平衡、表观消化率以及牛磺酸和羟脯氨酸等活性物质的含量上均具有优势,而且不含抗营养因子,传统上归为外因的差异。实践发现,尽管可以找到蛋白含量更高、表观消化率更优的蛋白源,也可以补充牛磺酸、氨基酸等,甚至可以去除抗营养因子,但是替代鱼粉后仍然无法达到预期的效果。研究发现,鱼粉与其他蛋白源相比,推动体蛋白沉积的能力存在显著差异,这就需要从内因角度研究,即从营养感知和蛋白质的代谢调控角度研究鱼体对不同蛋白源的反应。
借助于营养物质浓度梯度和鱼类生长的关系可获得最大生长速率时的营养需要量。研究发现,额外营养成分的添加对于鱼类生长的提高非常有限。据调查目前在实际养殖中,商品饲料都能够为养殖水产动物提供充足的营养,而且往往过营养化。如凡纳滨对虾的蛋白质需求在28%~32%,但有些饲料企业为了提高产品的“竞争力”,把对虾饲料蛋白质水平提高到了40%~42%。其实,多余的这部分蛋白质对对虾生长和蛋白质沉积的贡献非常小。深入剖析目前我国竞争激烈的饲料市场,几乎所有的产品都在追求高营养指标、高消化率,而忽视了由于营养过剩造成的水产动物生长、免疫和品质等下降的问题。这样带来的后果是增加了环境负荷和成本,又浪费了宝贵的蛋白源原料,这也是影响水产养殖业可持续发展的一个重要原因。实际上,未来水产动物营养研究更为关键的是将摄入的充足营养更多地用于机体蛋白质沉积,提高产出效率,让多余的营养素继续发挥作用,水产养殖动物持续增长。为了实现未来的研究目标,深入解析蛋白质被消化吸收以后的代谢和调控、决定饲料蛋白质沉积潜力的内在因素具有重要意义。
对高等动物与模式动物的研究证明,mTOR是调控体蛋白合成与生长的关键信号分子。mTOR可接收来自氨基酸、糖类、激素等各类外界信号,成为合成代谢的一个关键点。研究证实,鱼类饲料中添加mTOR抑制剂抑制其活性,将大大降低鱼体蛋白质的沉积,造成鱼类生长速度下降。这意味着饲料蛋白质即便被充分地消化吸收了,但并没有转化为体蛋白,即决定体蛋白沉积效率的开关是mTOR信号通路。鉴于此,如何激活鱼体内的mTOR信号通路,以及如何消除抑制mTOR信号通路的关键因子,对提高饲料蛋白质的利用率就显得尤为重要,是提高饲料蛋白质利用率的新思路。
研究发现,鱼类分别摄食鱼粉与其他蛋白源饲料后,其他蛋白源激活鱼体mTOR的效率显著不如鱼粉。饲用鱼粉蛋白饲料会产生一个高强度、持续时间较长的mTOR的激活,但饲用其他蛋白源饲料激活mTOR水平相对较低,这就必然造成鱼粉替代后鱼体蛋白合成不足,从而导致生长下降。虽然晶体氨基酸激活mTOR的强度也很高,但维持时间很短。40%的鱼粉被大豆蛋白取代后,在饲料中添加氨基酸,并没有显著改善体蛋白质的合成。由此可以得出,水产饲料中晶体氨基酸的添加并不能完全弥补鱼粉替代造成的mTOR信号通路激活和体蛋白质合成不足的问题。
mTOR作为调控体蛋白质合成的关键信号分子,氨基酸、抗营养因子、小分子活性物质均对mTOR活性有调节作用。平衡的氨基酸模式和小分子活性物质的存在有利于mTOR信号通路的激活,而氨基酸不平衡、抗营养因子的存在会抑制mTOR信号通路。例如饲料中常缺乏蛋氨酸,蛋氨酸的缺乏会抑制mTOR信号通路,降低新蛋白质的合成。棉粕中常见的抗营养因子——棉酚,也是通过抑制mTOR信号通路,从而抑制体蛋白质的合成。此外,所有的植物凝集素,如大豆凝集素、花生凝集素,都对mTOR有抑制作用,并会引起细胞凋亡。
研究发现,有一些物质可激活mTOR信号通路。mTOR的激活剂如磷脂酸能够显著提高细胞蛋白合成。当在细胞培养基中添加1~10 μmol/L的磷脂酸会激活并加强mTOR信号通路的表达,促进细胞分化和蛋白质合成。在大菱鲆饲料中添加0.5%磷脂酸,显著促进了体蛋白质的合成,鱼体终重提高了13.6%。β-羟基-β-甲基丁酸(HMB)也可以激活mTOR,添加0.1% HMB对鱼的特定生长率、肌肉品质都有明显的改善。
此外,投饲策略的差异也会显著影响mTOR的表达。投喂饲料后,经鱼体消化吸收后的氨基酸和其他营养物质能被mTOR信号通路感知,并反映在其活性的表达和体蛋白合成上。研究发现,当鱼摄食时mTOR表达上升,体蛋白质合成;鱼不摄食时mTOR不表达,体蛋白质就会分解。因此,mTOR的活性表达对氨基酸的摄入非常敏感。添加不同浓度的氨基酸时,肝脏和肌肉中的mTOR相关蛋白的表达具有显著差异。投喂频率对于mTOR的表达也有显著影响,比较研究1天投喂1次和1天投喂3次两种投饲策略,发现1天饲喂3次时激活mTOR的时间和强度要比1天投喂1次明显增加,鱼的生长明显改善。此外,通过增加投喂频率和添加激活剂——亮氨酸(1%)双重因素可大幅度增加mTOR活性,显著改善蛋白质的合成。
综上所述,为了有效提高饲料蛋白质利用效率,从内部因素考虑核心目标是激活mTOR信号通路和/或消除其抑制因素。
提高饲料蛋白质利用率的技术实践
提高饲料蛋白质利用率对保障水产养殖业可持续发展具有重要意义。针对mTOR信号通路,借助于整合氨基酸、活性肽、维生素、微量元素等能靶向激活调控mTOR等关键要素,开发了代谢信号复合调节包(Sig-Pep),在提高饲料蛋白质的利用效率上取得了一定成绩。以初重1.6 g的凡纳滨对虾为养殖对象设计饲喂试验,在山东胶州面积为238 m2的半土池,投苗约1万尾,每组3个池,盐度5‰,pH 8.0,水温26~30 ℃。将0.1% Sig-Pep与商品饲料拌料使用,每日投喂4次,连续饲喂一个月,试验结果见图1~图4。
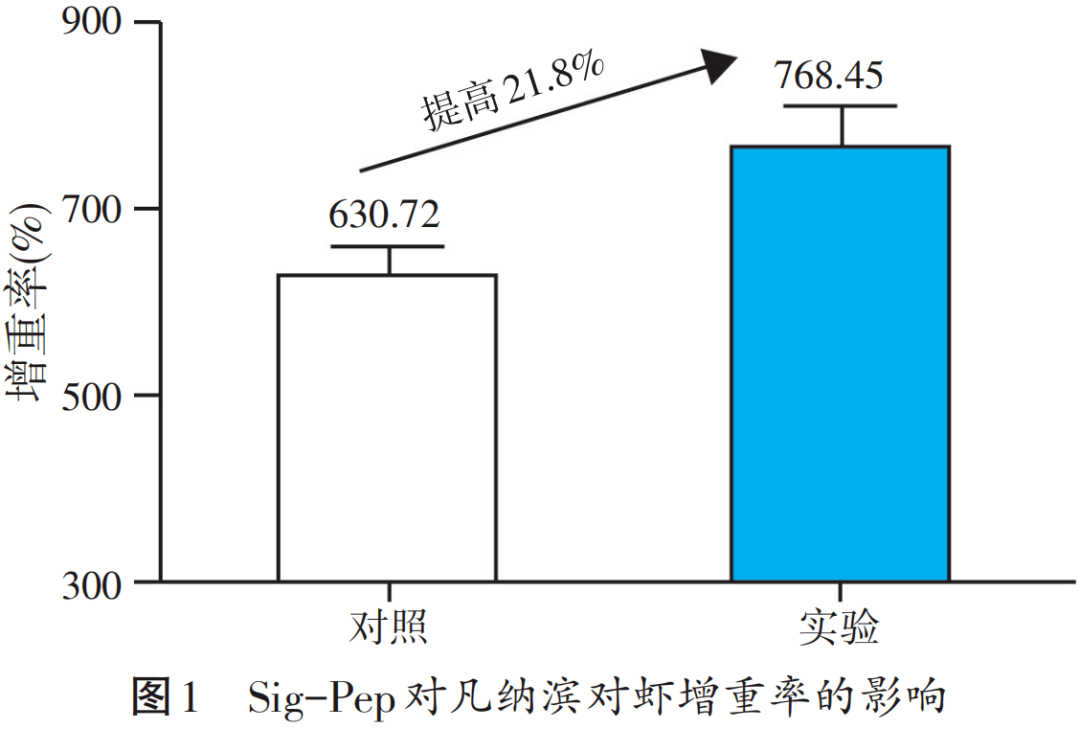
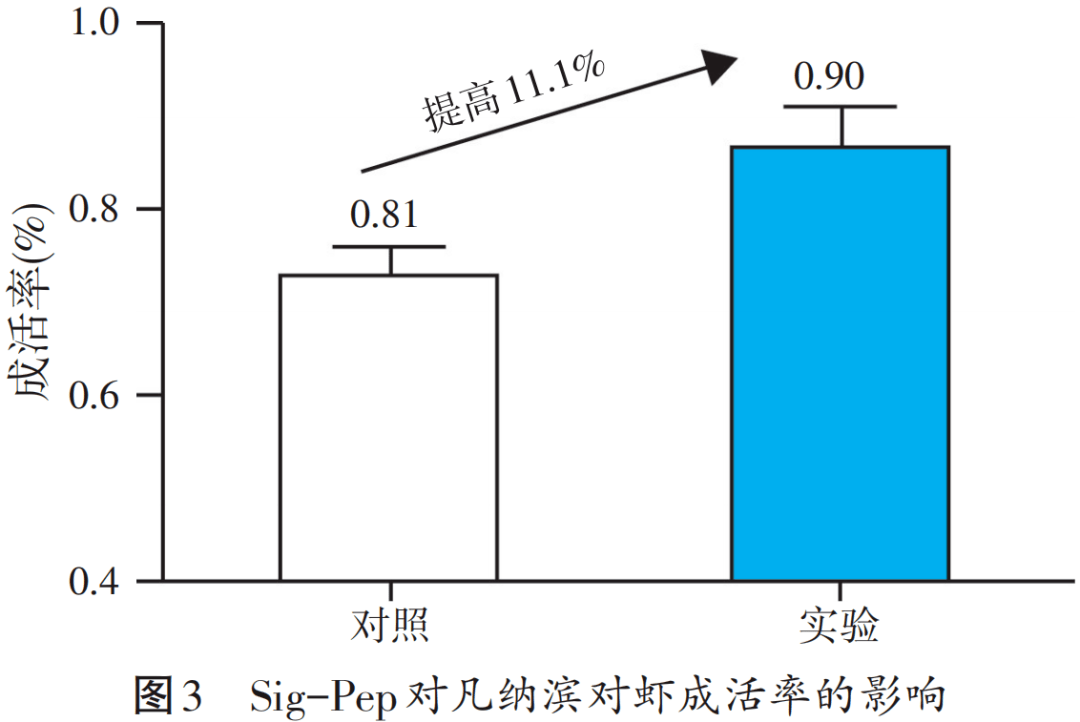
从图1~图4试验结果可见,Sig-Pep对凡纳滨对虾的增重率、饵料系数、成活率和产量均有显著效果,其中增重率提高了21.8%,饲料系数从0.99降到了0.84,成活率高达90%,每个池子的产量从101 kg增加到了123 kg。此外,Sig-Pep的添加可显著抑制对虾肝胰腺弧菌载量(见图5),显著改善非特异性免疫指标,显著提高了对虾的肥满度与虾青素含量(见图6)。结合凡纳滨对虾价格和饲料成本,据粗略统计添加量为1 kg Sig-Pep/t时,可增收7 200元,经济效益显著。
实践证明在饲料不用鱼粉或用植物蛋白替代大部分鱼粉时,添加Sig-Pep均能够促进水产动物的蛋白质合成和水产动物生长,从而减少鱼粉的使用。
Sig-Pep在鸡、猪等陆生动物上的应用研究同样取得了显著效益,旨在为无抗养殖提供技术。








